Recursion and Oscillations: Comments on Boeckx (2016) and Goucha et al. (2016)
4 responses to “Recursion and Oscillations: Comments on Boeckx (2016) and Goucha et al. (2016)”
-
Thanks Elliot for reading the paper and commenting on it. There are a few things I’d like to clarify, though, since you may have missed them.
You are quite right that there is an aspect of Friederici’s model that I find interesting, and that no doubt a lot of other people find interesting (including Chomsky and Berwick, which I mention in the paper, by the way). But the point of the paper was to stress the possibility of identifying traces of descent in this ‘special’ dorsal circuitry, and avoid “only us” approaches to language. In a sense, the slogan is ‘specialized, yes; special, no”.
You point out that “Pairing two finite-state machines does not produce a higher-order device” and seem to refer to a claim in my paper. But I address this point in the paper: I refer to works (such as Graf’s, or work I did with Uriagereka) stressing that we do not need to go beyond finite-state limits, so I am not claiming we necessarily need to produce a higher-order device (I think this is a very important point in the context of the paper, and the emphasis on decent).
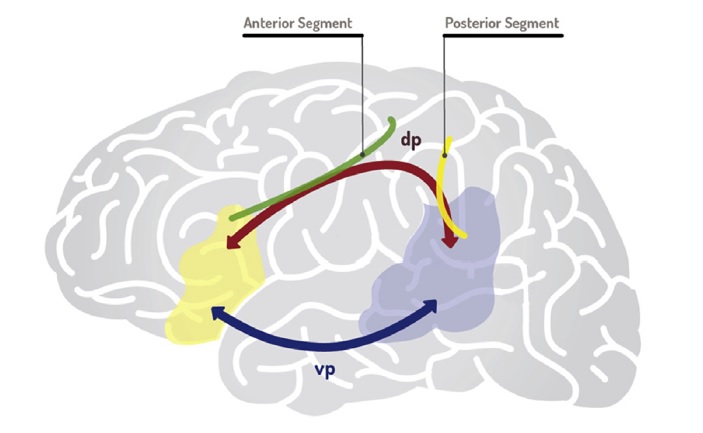
As for the observations about brain rhythms, you take issue, not with me, but with Goucha et al. I don’t address this point in the paper you blog about, but I should add that I side with Goucha et al. on this point: I think that cross-frequency couplings (theta-gamma, etc.) cannot be taken to be the source of human-specificity: they are the universal syntax of the mammalian brain, as Buzsaki would say. I suspect that connectivity plays a bigger role here: it’s not so much which frequencies get coupled with which ones, but rather which regions producing these frequencies can be interconnected. That’s why connectivity issues are central to the paper you comment on.-
Thanks Cedric. On the whole, I did find your paper interesting and certainly framed within the right kind of neuroanatomical context. I’ll reply to your comments in turn:
“(including Chomsky and Berwick, which I mention in the paper, by the way)”: My point was not that you didn’t cite the paper, since you do so even on the first page. Rather, I noted that the general “sequence pairing thesis” seems to be present in some form or another in Why Only Us (a book which I’m actually highly critical of).
I’m in full agreement that the correct slogan should be “specialized, yes; special, no”. In fact that’s exactly why I’m interested in exploring cross-frequency couplings which are well-known to be highly conserved. As the papers of mine cited discuss, it’s certainly not just the phase-amplitude theta/beta-gamma nesting algorithms which are relevant to language (since as my recent Theoretical Linguistics commentary article notes, non-human primates are also capable of these forms of cross-frequency relations) but rather *where* in the brain these “extraction” and “maintenance” operations are taking place. So I’m agreement on your point about connectivity being central.
When it comes to the finite-state issue, I did indeed notice your alignments with Graf et al in the paper, but my main issue was that the jump from “sequences” to “sequences of sequences” upon the ventral-dorsal pairing (again, as claimed in Why Only Us) would have to be given some kind of independent neurocomputational/oscillomic motive. So, going back to brain rhythms, this will likely be a nice way to blend the findings of the neuroimaging “cartographic” literature with the oscillomic work on attention, semantic memory, and phrase structure building. Of course there’s nothing special about hippocampal theta and thalamic alpha, but upon the emergence of a more globularised braincase (as you, and Antonio and myself have discussed recently) the generic extraction/maintenance operations of, for instance, the theta-gamma code would be able to “reach” into new areas previously cordoned off, so to speak (eliminating some of those “spatial inequalities” found in non-human primate brains).-
Ok … so it seems you disagree with me far less than your original comment(‘s rhetoric) suggested. Good to know. I note in the 1st footnote of the paper that the proposal I make could relate to the one by Chomsky and Berwick, though I doubt it, because if it did, it would defeat their “why only us” point. The “sequence of sequence” step is argued for in my paper in part on the basis of the _hierarchical_ relation between Dehaene’s model and more modular networks (cf. my point about sequences of chunks). I still think you miss the point about ‘finite-state’, and it seems there is another point you miss: you mention “ventral-dorsal”, but I think one of the important points is “dorsal I and dorsal II” (Friederici’s insight, in my view). “Ventral” plays no role in what I discuss (i.e., I do not “jump from “sequences” to “sequences of sequences” upon the ventral-dorsal pairing”). Finally, if you agree with me about connectivity, not sure what the big deal about oscillation was in the original blog post. Anyway, thanks for reading the piece, and taking the time to comment, Elliot.
-
My intention was not to point to a clear disagreement with your paper so much as it was to draw attention to current hurdles in investigating (and providing support for) its central claims. I get your reservations about the “only us” part of Why Only Us (something I share), but again that wasn’t what I was really interested in – it was more to do with the fact that the general loop-based architecture had been proposed in the book (and in some other places, too) and I found it odd that since you already cite the book in the paper you didn’t also note this similarity. I also get the point about the pairing of finite-state sequence processors, and in fact I got it straight away when I read the paper. As I said, I was puzzled by the sudden “jump” implied by pairing two sequence processors: How, in implementational/algorithmic terms, would the streams embed finite-state sequence X with finite-state sequence Y, yielding “sequences of sequences”? Finally, there’s no “big deal” about oscillations in the original post, it’s only a single paragraph and I was more interested in pointing to the (current) lack of a story here w/r/t linking the oscillomic code (developed in my own work) with the neuroanatomical novelties pointed to in Why Only Us, Friederici’s work, and much else (including the Theoretical Linguistics commentary article I mentioned).
-
-

Leave a comment